気孔孔辺細胞は青色光に反応してH+を放出し気孔を開かせる
解明された気孔の青色光受容体
理学研究院・生物科学部門 教授 島崎研一郎
今回は、理学研究院の島崎研一郎教授が、植物における光情報に対する応答機能研究の最前線を、気孔をモデルとして分かりやすくご紹介します。なおこの研究は、新たな学問領域の創設や教育の推進を目的とする本学独自の制度「教育研究プログラム・研究拠点形成プロジェクト」(通称P&P)に、平成11-12年度に採択されたものの一部です。島崎教授を代表とする研究組織に加わったのは、理学研究院、農学研究院でした。
〈はじめに〉
植物葉の気孔は光合成の材料になる炭酸ガスを取り入れるとともに、根から木の頂までの水の吸い上げを可能にする、植物にとって大変重要な器官です。気孔のない植物も例外的に存在しますが、アンデスの高地ペルーで発見されたものは、炭酸ガスを土壌中から得ており、水を吸い上げる事ができないので、地面に張り付いて細々と生活しています。高等植物の気孔は光に反応し、特に、青色の光に敏感に反応して開口する事から、気孔の青色光反応と呼ばれています。植物の青色光反応には、進化論で知られるC・ダーウィンの研究で知られる光屈性をはじめとして、モヤシ化を防ぎ緑化させる光伸長抑制、カロチノイドの生合成反応、呼吸の促進など多くのものがあり、大変長い研究の歴史を持ちますが、それらの反応の初発過程を担う青色光受容体が解明されたのはごく最近の事です。1993年から2001年にかけて、光伸長抑制、光屈性、葉緑体運動などの青色光受容体が次々と解明されるなかで、気孔の青色光受容体は最後に残された大きな課題でした。私達はこの気孔の青色光受容体を解明することが出来たので紹介します。
〈気孔はどのようにして開くか〉
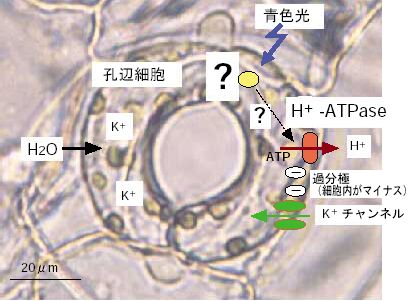 |
| 図1 青色光による気孔開口 |
光の当たっている植物葉を切り取ってきて机の上に置いておくと、そのうち萎れてしまいます。しかし、同じ植物葉を光が当たらないようにして、切り取ってもなかなか萎れません。これは、表皮に存在する気孔が光によって開いており、そこから水が蒸発(蒸散)するからです。暗中では気孔は閉じているのです。また、日中の木陰が涼しい理由の1つは蒸散によって葉の表面から熱が奪われるためです。このように、気孔は光が当たると開口しますが、この現象はC・ダーウィンの子フランシス・ダーウィンが記述しており、彼は、窓側に近い気孔ほど良く開いていることに気がつきました。青色光はどのようにして気孔を開かせるのでしょうか?気孔開口はこの器官を構成する一対の孔辺細胞に高濃度のK+が蓄積し(数百ミリモルに達する)、その浸透圧増加に伴い水が取り込まれ、細胞の体積が増加するためである事が分かっています。私達は1986年に青色光が孔辺細胞の細胞膜にあるプロトンポンプを活性化し、それによってH+を細胞の外に電位差形成的に輸送し、孔辺細胞の膜電位を過分極(よりマイナス側へ電位をシフトさせる)させる事を示しました(図1)。その翌年、パッチ・クランプ法の創始者の一人E.Neherも参加した研究によって、孔辺細胞の細胞膜に膜電位に依存して開口する内向き整流性のK+チャネルが見出され、このチャネルは孔辺細胞の膜電位が-120mV以下になると開口する事が証明されました。従って、青色光は細胞膜プロトンポンプを活性化し、膜電位を-120mV以下に過分極させ、過分極に依存して活性化されるK+チャネルを通して外液のK+を濃度勾配に逆らって取り込み、蓄積させる事になります(図1)。私達は最近、このプロトンポンプの実体が細胞膜H+-ATPaseであり、この酵素のC-末のスレオニンが青色光によってリン酸化され、その部位にいわゆる14-3-3タンパク質が結合して活性化される事を明らかにしました。そこで、青色光を吸収し、化学シグナルに変換してポンプに伝える青色光受容体がどのような物質か、大変興味ある問題です。いくつかの説が出されましたが、いずれも多くの研究者の納得を得るには至らず、問題が残されたままでした。
〈気孔の青色光受容体は何か〉
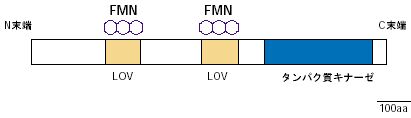 |
| 図2 photの基本構造 |
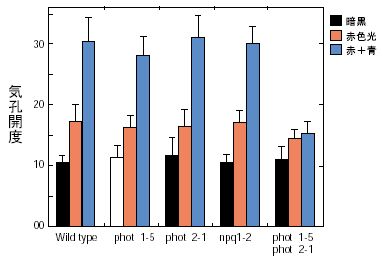 |
| 図3 シロイヌナズナ変異株における光による気孔開口 |
1997年になって、米国のグループによって青色光を照射しても光屈性を示さない変異株が単離されました。この変異株は生化学的研究によって青色光受容体を欠いている事が分かってきましたので、それを手がかりに青色光受容体をコードする遺伝子が同定され、そのタンパク質はフォトトロピズム(光屈性)にちなんでフォトトロピンと名付けられました。フォトトロピンは、N末に青色光受容色素として2個のフラビンモノヌクレオチド(FMN)が結合しており、C末部のプロテインキナーゼと融合した受容体型キナーゼタンパク質でした(図2)。このタンパク質の吸収スペクトルは気孔開口の作用スペクトルと大変良く似ており、気孔の青色光受容体である可能性があります。しかし、フォトトロピンを欠いた変異株は青色光照射によって気孔が開口したので、これとは別に気孔の青色光受容体が存在すると考えられました。ところが、フォトトロピンと良く似た構造を持つ新たな青色光受容体が発見され、2001年3月にこの物質が強光から逃れる葉緑体運動に関与する事が示され、最初のものはphot1、葉緑体運動に関与するものはphot2と名付けられました。phot1とphot2は良く似た吸収スペクトルを示し、ひょっとしたらこの二つの青色光受容体の両方とも気孔開口に関与するのかも知れません。もしそうなら、どちらか片方の青色光受容体が無くても、残りが働いて青色光を当てると気孔が開く事が考えられますし、今まで多くの研究者が変異株のスクリーニングをくり返しても気孔の変異体が得られなかった理由が説明出来ます。この事を確かめるにはphot1とphot2の両タンパク質を欠いた2重変異株を作出して実験を行う必要があります。ちょうどそのころ基礎生物学研究所の和田研究室でこの2重変異株を得ており、早速、これを用いて気孔の反応を調べるとphot1、phot2の単一変異株の気孔は青色光に反応してそれぞれ開口するのに、phot1、phot2の二重変異株だけが全く反応しませんでした(図3)。また、この二重変異株のみが青色光によるプロトン放出も完全に阻害されました。さらに、従来、青色光受容色素としてカロチノイドの一種ゼアキサンチンが提案されていましたが、この色素を合成できないnpq変異株の気孔も青色光で開口しました(図3)。これは、やはりphot1とphot2が重複して気孔開口を引き起こす事を示しています。ただし、二重変異株は気孔開口の駆動力となるプロトンポンプ、あるいは、内向き整流性K+チャネルが傷んでいる可能性があります。しかし、プロトンポンプの含量、活性、あるいはK+チャネルの働きは野生株と差が無く、青色光に対して気孔が反応しないのは青色光受容体phot1とphot2の両方が欠けているせいである事が確証されました。この成果は、植物生理学上の長年の問題を解決した論文として公表しました(Kinoshita et al.,Nature 414: 656-660, 2001)。気孔が光に反応する事がF・ダーウィンに見出されてから100年以上経過していました。毎朝、陽が昇ると、地上の樹木、草木、作物の葉が、この二つの青色光受容体に吸収された光に反応して孔辺細胞からH+を放出させ、気孔を開かせていると想像するのは楽しい事です。
(しまざき けんいちろう 植物細胞機能学)
前のページ
ページTOPへ
次のページ
インデックスへ